III) La reproduction sexuée brasse les allèles au sein de la population
A) La reproduction sexuée est caractérisée par deux événements fondamentaux : la méiose et la fécondation
Au cours de la reproduction sexuée, un gamète mâle, très souvent mobile, féconde un gamète femelle, le plus souvent immobile et riche en réserves cytoplasmiques. Chacun des deux gamètes, haploïde, apporte n chromosomes (23 chez l’Homme), la cellule-oeuf qui résulte de la fécondation de l’ovule par le spermatozoïde possédant alors 2n chromosomes (46 chez l’Homme). Le mélange du contenu chromosomique des deux noyaux gamétiques est appelé la caryogamie.
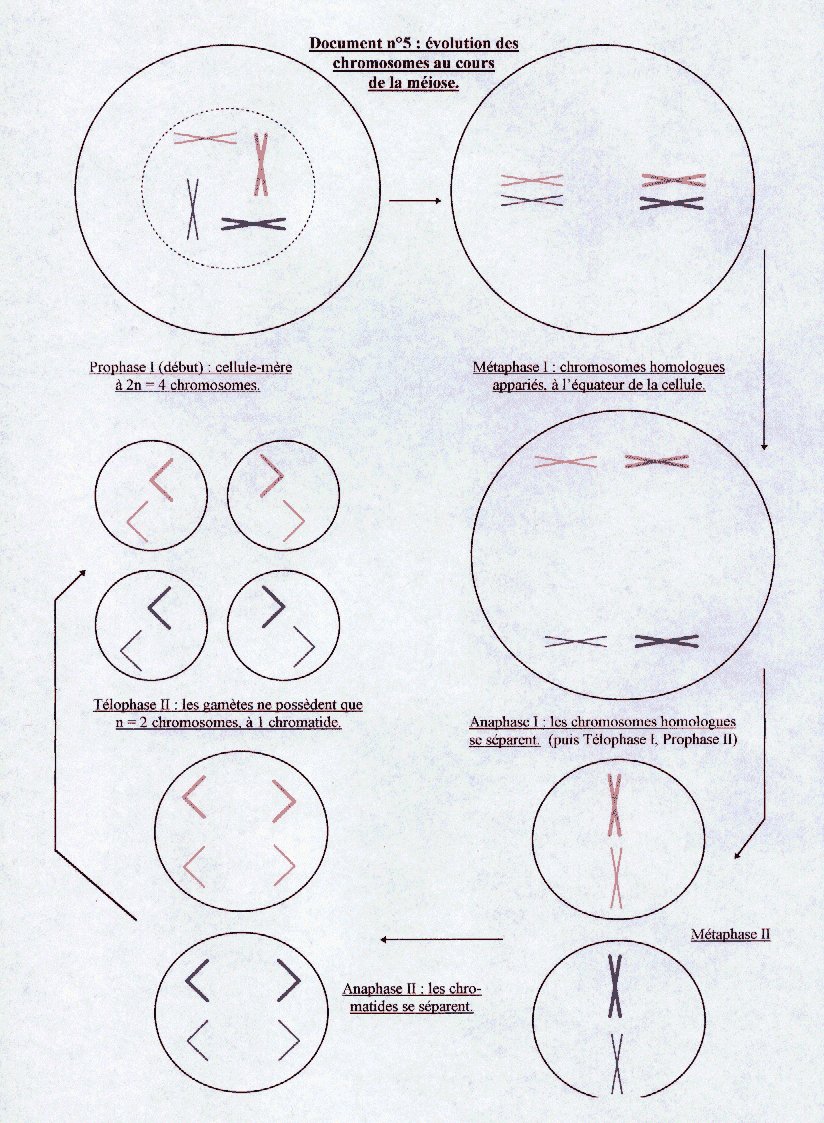
Les cellules haploïdes sont obtenues par méiose à partir de cellules diploïdes. La méiose est composée de deux divisions successives, sans synthèse d’ADN entre les deux. Au cours de la première division de la méiose, en prophase I, les 2n chromosomes se condensent puis ils s’apparient : on obtient n paires de chromosomes homologues étroitement accolés sur toute leur longueur. Pendant cette étape, les deux chromosomes forment des figures de chiasma : les chromatides homologues sont entrecroisées, et des segments homologues de chromatides peuvent être échangés. Ces échanges sont appelés crossing-over. En métaphase I, les chromosomes appariés, en partie recombinés, se placent sur le plan équatorial de la cellule. En anaphase I, les deux chromosomes homologues de chaque paire se séparent, et migrent chacun vers un pôle opposé de la cellule. Aux deux pôles de la cellule se retrouvent alors n chromosomes à deux chromatides. Il s’ensuit une division qui ressemble à une mitose, les quatre cellules obtenues n’ayant plus que n chromosomes à une chromatide. ( voir document n°5)
Suivant la place respective de la méiose et de la fécondation, dont la caryogamie, on distingue de nombreux cycles biologiques de développement. Les plus simples se rencontrent chez les organismes diploïdes (l’Homme par exemple), où la fécondation suit immédiatement la méiose, et chez les organismes haploïdes (les champignons Sordaria par exemple), où la méiose suit immédiatement la fécondation.
B) Un individu produit des gamètes parmi une infinité de possibles
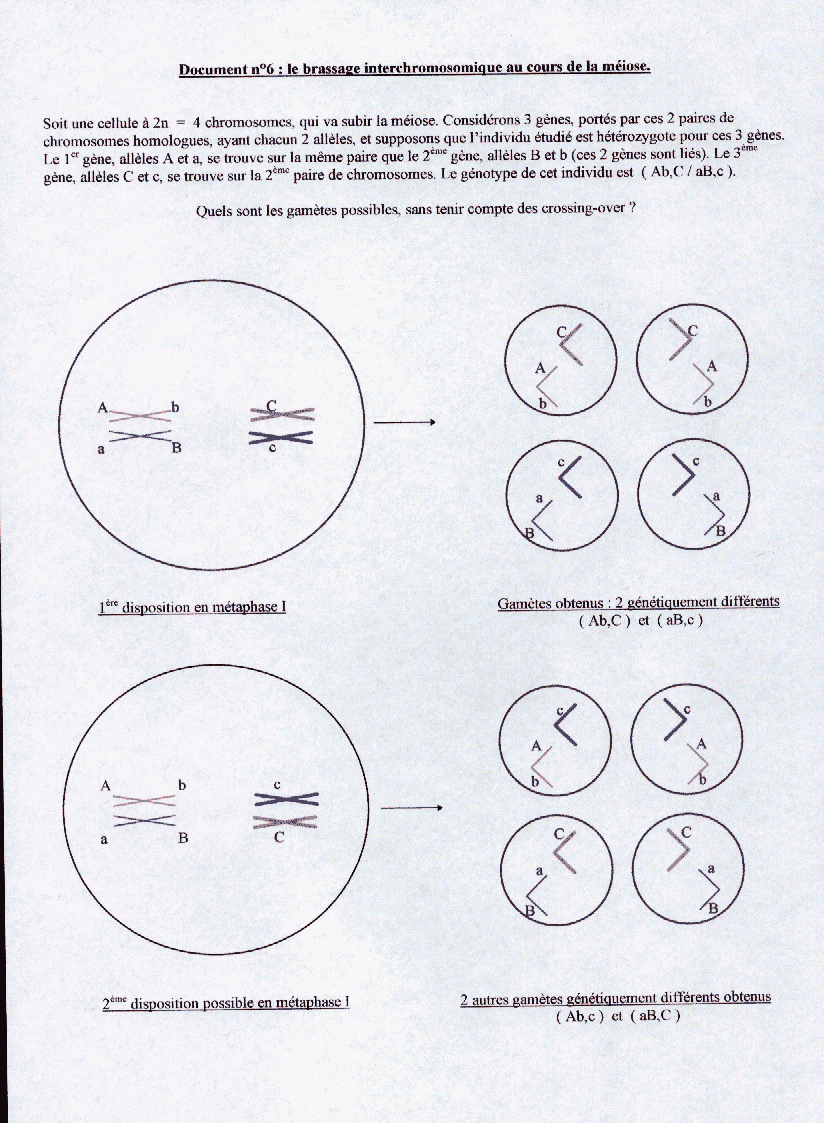
En métaphase I de méiose, les deux chromosomes homologues de chaque paire restent l’un près de l’autre, et ils se placent à l’équateur de la cellule, un chromosome d’un côté du plan équatorial, l’autre chromosome de l’autre côté, et lorsqu’ils se sépareront, en anaphase I, chacun migrera vers le pôle le plus proche. Or on constate, statistiquement, que le hasard intervient dans le placement de ces chromosomes : chaque chromosome de chaque paire se positionne d’un côté ou de l’autre au hasard. Il en résulte que les cellules d’un individu, lorsqu’elles se divisent par méiose, « choisissent » une métaphase I parmi un grand nombre de métaphases I possibles. Un individu pourra alors produire un grand nombre de gamètes différents par les chromosomes qu’ils contiennent. Ce nombre sera d’autant plus grand que le nombre de paires de chromosomes homologues est élevé, soit 2n gamètes possibles. Pour l’Homme, n = 23 et 223 = 8 388 608 !
Ce brassage génétique est appelé brassage interchromosomique. ( voir document n°6)
En prophase I, les chromosomes homologues de chaque paire ont pu échanger un segment de chromatide, plus ou moins long. Ces crossing-over se produisent là encore au hasard. Le brassage interchromosomique affecte des chromosomes en partie recombinés par un brassage intrachromosomique. Comme la longueur du segment échangé semble dépendre du hasard, tout se passe comme si chaque gène pouvait subir une recombinaison au sein de chaque paire. Ces crossing-over modifieront la combinaison allélique d’un chromosome seulement si les deux allèles du gène concerné sont différents, c’est à dire si l’individu est hétérozygote pour le gène concerné. ( voir document n°7)
Chez l’Homme, on peut estimer à quelques % la quantité de gènes pour lesquels un individu est hétérozygote, soit environ une centaine de gènes sur chaque chromosome (le nombre de gènes sur les 46 chromosomes est d’environ 100 000). Le nombre théorique de gamètes génétiquement différents qu’il est alors possible de produire à partir d’une seule métaphase I est de 2100.
En tenant compte des deux brassages génétiques successifs, un individu humain peut théoriquement produire environ 2 23 x 2 100 = 2 2300 gamètes génétiquement différents. C’est à dire que chaque gamète produit (spermatozoïde ou ovule) contient une combinaison allélique parmi une infinité de combinaisons possibles.
C) Chaque individu obtenu par reproduction sexuée est unique
La fécondation accentue la diversité génétique des descendants possibles d’un couple. En effet, un spermatozoïde parmi une infinité de possibles va féconder un ovule parmi une infinité de possibles. Pour reprendre l’exemple de l’espèce humaine, le nombre d’assortiments alléliques différents qu’un couple peut engendrer est alors de 2 2300 x 2 2300 soit 2 4600. Il est donc impossible, à l’exception des vrais jumeaux, que deux frères et soeurs soient génétiquement identiques. De même on peut affirmer sans crainte que chaque être humain porte une combinaison allélique originale : jamais personne n’a possédé ni ne possédera la même combinaison d’allèles. Langaney résume cette idée ainsi : « qui fait un oeuf fait du neuf ».
Conclusion : la reproduction sexuée est un mode de reproduction adapté lorsqu’il faut une grande diversité de phénotypes dans la population, sur lesquels va s’exercer la sélection naturelle. Ceci rentre bien dans la théorie darwinienne de l’évolution : la sélection naturelle opère un tri, une pression sur les individus de la population, tous différents lorsqu’ils sont obtenus par reproduction sexuée, car possédant chacun une combinaison originale d’allèles. Cependant la méiose et la fécondation ne créent pas de nouveaux allèles, ces derniers sont obtenus par mutations.
D) La méiose peut jouer un rôle « réparateur »
Si pour une minorité de gènes, le polyallélisme est important dans la population, et est utile du point de vue de l’évolution, pour la plupart de nos gènes, peut-être 90 %, il n’existe qu’un allèle, et nous sommes homozygotes. Sans doute que pour la majorité de ces gènes, la variabilité n’est pas bénéfique, et il convient de les conserver inchangés. La recombinaison génétique observée au cours de la méiose, les crossing-over, permettent ainsi d’éliminer des mutations, des anomalies, tout en restaurant, dans certains gamètes, la bonne combinaison allélique, celle qui est inchangée.
Exemple : Dans le cas d’un individu de génotype ( Ab,C / aB,C ) , en supposant que les 2 allèles a et b sont désavantageux, un crossing-over peut rétablir la « bonne » combinaison allélique, exprimée dans les gamètes ayant pour génotype ( AB,C ).
L’importance de ce rôle « réparateur » de la méiose reste à approfondir, mais on ne peut pas l’exclure à priori, même s’il paraît moins évident que son rôle de brassage génétique au sein de la population. Réexaminons le cas des organismes eucaryotes, tels que les pucerons, qui ont recours à la reproduction sexuée de temps en temps, et qui prolifèrent habituellement par reproduction asexuée, par parthénogenèse par exemple. Un inconvénient de la reproduction asexuée, c’est le risque d’accumulation de mutations néfastes, traînées par les individus comme un fardeau de plus en plus lourd à chaque génération. La reproduction sexuée, grâce à la recombinaison génétique entre chromosomes homologues, peut permettre de revenir à des individus sans mutations. En effet, deux parents atteints de mutations différentes peuvent donner naissance à des descendants sans mutations.
Remarque : en éthologie, la reproduction sexuée chez les Primates a été particulièrement étudiée. Il a été montré, chez les Bonobos, que différents comportements sexuels jouent un rôle d’apaisement des tensions au sein du groupe, accompagnés parfois d’un partage de nourriture. (Voir La Recherche n° 210, de mai 1989, page 597, et Pour la Science n° 211, de mai 1995, page 70)
E) Les inconvénients de la reproduction sexuée
Le « coût de la production des mâles » représente un inconvénient majeur ( voir document n°8). Une population s’accroît beaucoup plus vite à partir de femelles parthénogénétiques qu’à partir de femelles ayant recours à la reproduction sexuée. D’une façon générale, le coût énergétique de la reproduction sexuée ne plaide pas en sa faveur. On constate ainsi, chez les espèces où la rencontre entre les deux sexes est difficile, des adaptations pour surmonter cette contrainte. Citons le cas de poissons abyssaux, où le mâle est réduit, de très petite taille, et fixé comme un parasite sur le corps de la femelle (voir La Recherche n° 281, de novembre 1995, page 44). Chez certains organismes parasites, le corps de la femelle est comme greffé à celui du mâle.
Remarque : le cas des individus hermaphrodites. A la fois mâles et femelles, les hermaphrodites ont souvent recours à une fécondation croisée, comme chez les escargots. Chaque individu va alors pondre des oeufs. L’observation du comportement d’un ver marin hermaphrodite, Pseudoceros bifurcus, montre le « coût de la féminité » : produire des oeufs nécessite une grosse dépense d’énergie, un coût important, que chacun des deux vers, pendant l’accouplement, semble vouloir éviter. Ils se font face, cherchant à planter leur pénis dans le corps de l’autre, et à y injecter leur sperme, tout en esquivant les tentatives d’insémination traumatique. Celui qui « gagnera » transmettra ses allèles à ses descendants, sans supporter ce coût de la féminité. (voir la Recherche n° 324, page 44)
Du point de vue du gène, son « succès » évolutif, à court terme tout au moins, est meilleur par la reproduction asexuée, car un organisme lègue alors la totalité de ses allèles à ses descendants, contre la moitié seulement par la reproduction sexuée (voir la théorie du gène égoïste, pour laquelle les organismes ne sont que des artifices empruntés par les gènes pour se reproduire).
Si la reproduction sexuée prévaut largement, elle doit être efficace à court terme. Or lui trouver des avantages à court terme n’est pas évident, si ce n’est peut-être la nécessité d’évoluer sans cesse, pour supporter la pression de sélection imposée par les facteurs biotiques de l’environnement : autres espèces concurrentes, parasites, agents pathogènes... C’est l’hypothèse de la Reine rouge (L. Van Valen) : cf. la reine rouge, de Lewis Carroll, qui force Alice à courir aussi vite qu’elle le peut pour rester à la même place. Citons aussi ces expressions : « l’enfer, c’est les autres », et « qui n’avance pas recule ».
En fait, le recours périodique à la reproduction sexuée chez les espèces se reproduisant par voie asexuée, s’il ne peut pas s’expliquer facilement comme étant une adaptation, peut mieux se comprendre comme étant une exaptation, c’est à dire un caractère hérité de lointains ancêtres, à une période où ce caractère était avantageux. Cela peut être la présence de structures de dispersion des jeunes ou des graines obtenus par reproduction sexuée (ailes des pucerons non parthénogénétiques, dispersion des graines à longue distance grâce à des structures adaptées...), ou la résistance au froid des oeufs non parthénogénétiques.
Conclusion : Tout conduit à penser que la reproduction sexuée s’est imposée, et se rencontre massivement chez les espèces eucaryotes actuelles, car les espèces ayant perdu cette capacité ont été éliminées. Elles n’ont pas pu franchir, par exemple, les crises d’extinction passées. En somme, nous avons hérité d’une reproduction sexuée car nos lointains ancêtres la possédaient.
BILAN : apparemment, la reproduction sexuée l’emporte nettement sur la reproduction asexuée, tout au moins pour les organismes eucaryotes. Ainsi, le mode de reproduction sexuée est plus efficace pour permettre aux espèces de franchir les périodes critiques, pendant lesquelles il est nécessaire d’évoluer rapidement.
Exemple : supposons qu’il faut posséder 3 mutations m1, m2, m3, pour être plus performant, mieux adapté au milieu. Dans une population où les individus se reproduisent par reproduction asexuée, ces mutations devront apparaître successivement, par exemple d’abord m1, puis m2 chez des individus mutés m1, puis m3 chez des individus mutés pour m1 et pour m2, ce qui prendra beaucoup de temps. Dans une population où les individus se reproduisent par reproduction sexuée, ces 3 mutations peuvent survenir chez des individus différents, à la même époque. Puis, par croisement, on pourra obtenir d’abord des individus possédant 2 mutations, et ensuite par d’autres croisements, des individus possédant les 3 mutations. On comprend aisément qu’il faudra alors moins de temps dans ce 2ème cas.
Problème : chez les procaryotes, la reproduction sexuée précédemment décrite n’existe pas. Or ils sont présents sur Terre depuis plus de 3,5 milliards d’années semble-t-il, ils sont extrêmement diversifiés et très bien adaptés à beaucoup de niches écologiques. De nombreux groupes de procaryotes ont ainsi franchi avec succès les crises biologiques, assez fréquentes au cours des temps géologiques, ce qui prouve leur efficacité, leur haut degré évolutif. Comment expliquer ce succès ?
{kind=link}
{kind=link}
{kind=link}
{kind=link}